
Nombres:
Starlin Galvan Minier 1-11-1168
Gladisnil Silverio Franco 1-12-1176
Ricardo jean 1-12-1137
Rose Darline
Francois 2-12-0770
Wideline Yeudy
2-11-1618
Noel Belmondge
1-12-2822
Rose Annie Marcellus
2-12-1074
Stanley Edouard
1-12-1340
Mirlanda Augustin
1-12-2655
Gabrielle Nohemie Chery
1-12-6227
Sony Pierre 1-12-1825
Jephte Wherley Rene
1-12-1816
Jinet Jaquez 2-11-1526
Stephanie Gil 2-09-0962
Temas:
Citoquinas, Inflamación y Fagocitosis.
Profesora:
Dra. Mirtha Villar
Grupo:
09
Asignatura: Inmunología
INTRODUCCIÓN
Citoquinas:
El desarrollo de una respuesta inmune efectiva implica a
diferentes grupos celulares. Las complejas interacciones entre estas células
están mediadas por una serie de proteínas secretadas de bajo peso molecular que
son conocidas colectivamente como citoquinas. Las citoquinas funcionan como
mensajeros del sistema inmune: regulan la intensidad y duración de la respuesta
inmune, estimulando o inhibiendo la proliferación de varias células, la
secreción de anticuerpos o de otras citoquinas.
Fagocitosis:
Este es un tema muy sencillo por el cual las
células blancas nos ayudan protegiéndonos de organismos a veces patógenos que
nos atacan.Antes que nada empezaremos con una sencilla pregunta:
Es una función de células
especializadas del sistema inmune, capaces de remover cuerpos extraños y
combatir infecciones del sistema inmune como primera linea de defensa natural.
Inflamación:
La inflamación es una
respuesta protectora cuyo fin es librar al organismo de la causa inicial de la
lesión celular (microorganismos patógenos, toxinas) y de las consecuencias de
la misma (células y restos tisulares necróticos). Sin el proceso de
inflamación, las infecciones se propagarían de forma incontrolada, las heridas
no se curarían nunca y los órganos lesionados presentarían lesiones supurativas
de forma permanente. No obstante, los procesos de inflamación y reparación
pueden ser perjudiciales: las reacciones inflamatorias constituyen el mecanismo
patogénico básico de las reacciones de hipersensibilidad potencialmente
mortales secundarias al efecto de picaduras de insectos, fármacos o toxinas y
también lo son de algunas de las enfermedades crónicas frecuentes (artritis
reumatoide, aterosclerosis, fibrosis pulmonar). La reparación mediante fibrosis
puede dar lugar a la aparición de cicatrices con desfiguración, y también a
bandas de fibrosis que producen obstrucción intestinal o limitación de la
movilidad articular. Los fármacos antiinflamatorios potencian los efectos
beneficiosos de la inflamación controlando al mismo tiempo sus secuelas
nocivas.
La citoquinas
Las citoquinas (o
citosinas) son un grupo de proteínas de bajo peso molecular que actúan mediando
interacciones complejas entre células de linfoides, células inflamatorias y
células hematopoyéticas.
Sus funciones son muy variadas, pero se pueden clasificar en unas pocas categorías:
diferenciación y maduración de células del sistema inmunitario;
comunicación entre células del sistema inmunitario;
en algunos casos, ejercen funciones efectoras directas.
En el pasado reciente hubo un cierto galimatías con la cuestión de su denominación. Así, muchas de las primeras citoquinas se descubrieron como señalizadoras entre leucocitos, por lo que se denominaron interleuquinas; otras eran secretadas por monocitos/macrófagos, por lo que se llamaron monoquinas. Sin embargo, muchas de esas sustancias son producidas por otros tipos celulares, por lo que se desaconseja el uso de esas denominaciones, para agruparlas a todas bajo el concepto de citoquinas. Las quimioquinas (o quimiocinas) son un tipo de citoquinas de pequeño tamaño, con papeles en la respuesta inflamatoria y la quimiotaxis de fagocitos.
14.2 PROPIEDADES GENERALES DE LAS CITOQUINAS
Las citoquinas son un grupo de proteínas secretadas de bajo peso molecular (por lo general menos de 30 kDa), producidas durante las respuestas inmunes natural y específica. Se unen a receptores específicos de la membrana de las células donde van a ejercer su función, iniciando una cascada de transducción intracelular de señal que altera el patrón de expresión génica, de modo que esas células diana producen una determinada respuesta biológica.
Las citoquinas son producidas por múltiples tipos celulares, principalmente del sistema inmune. Dentro del sistema inmune natural, los macrófagos son de las células más productoras de citoquinas, mientras que en el sistema específico lo son las células T colaboradoras.
La producción de las citoquinas suele ser breve (transitoria), limitada al lapso de tiempo que dura el estímulo (es decir, el agente extraño). En muchos casos ello se debe a que los correspondientes ARNm tienen una corta vida media, que a su vez depende de que las zonas 3’ no traducibles son ricas en A y U.
Considerando las diversas citoquinas, éstas pueden exhibir una o varias de las siguientes cualidades:
- pleiotropía (múltiples efectos al actuar sobre diferentes células).
- redundancia (varias citoquinas pueden ejercer el mismo efecto).
- sinergismo (dos o más citoquinas producen un efecto que se potencia mutuamente).
Por ejemplo, la acción conjunta de IL-4 e IL-5 induce en células B el cambio de clase para que produzcan IgE.
- antagonismo (inhibición o bloqueo mutuo de sus efectos).
Por ejemplo, el IFN-g bloquea el cambio de clase promovido por IL-4.
Las citoquinas ejercen su acción al unirse a receptores específicos para cada citoquina en la superficie de la célula en la que ejercen el efecto. La afinidad de cada receptor hacia su citoquina correspondiente suele ser bastante alta, del orden de lo femtomolar (10-15 M) a lo picomolar (10-12 M).
Utilizando la analogía de lo que ocurre con las hormonas del sistema endocrino, las acción de las citoquinas se puede clasificar en:
- de tipo autocrino
- de tipo paracrino
- (en pocas ocasiones) de tipo endocrino.
Las citoquinas "controlan" el sistema inmune de varias maneras, que podemos agrupar de la siguiente manera:
regulando (activando o inhibiendo) la activación, proliferación y diferenciación de varios tipos de células;
regulando la secreción de anticuerpos y de otras citoquinas.
14.3 ESTRUCTURA Y FUNCIÓN DE LAS CITOQUINAS
Las citoquinas son proteínas o glucoproteínas de menos de 30 kDa. Muchas de ellas pertenecen a la llamada familia de las hematopoyetinas, y tienen estructuras terciarias parecidas: una configuración a base de un conjunto de cuatro hélices a , con poca estructura en lámina b .
Generalmente actúan como mensajeros intercelulares que suelen intervenir en la maduración y amplificación de la respuesta inmune, provocando múltiples actividades biológicas una vez que se unen a los receptores específicos de las células diana adecuadas.
Aunque existen muchos tipos de células productoras citoquinas (ya hemos ido viendo unas cuantas en los temas anteriores), los más importantes son los linfocitos TH y los macrófagos, ya que sus citoquinas son esenciales para que se produzca la respuesta inmune una vez que se activan las células T y B por el contacto con las correspondientes células presentadoras de antígeno.
Principales tipos de respuesta mediatizados por la acción de las citoquinas:
activación de los mecanismos de inmunidad natural:
activación de los macrófagos y otros fagocitos
activación de las células NK
activación de los eosinófilos
inducción de las proteínas de fase aguda en el hígado (ver tema 16, sobre el complemento).
Activación y proliferación de células B, hasta su diferenciación a células plasmáticas secretoras de anticuerpos.
Intervención en la respuesta celular específica.
Intervención en la reacción de inflamación, tanto aguda como crónica.
Control de los procesos hematopoyéticos de la médula ósea.
Inducción de la curación de las heridas.
Hay una aparente paradoja de las citoquinas que debemos explicar: ¿Por qué las citoquinas, que son inespecíficas respecto del antígeno, pueden ejercer acciones de modo específico? Veamos varios mecanismos:
Regulación muy fina de los receptores de cada citoquina: los receptores celulares indispensables para que una citoquina ejerza su papel sólo se expresan en tipos celulares concretos una vez que éstos han interaccionado con el antígeno (pensemos por ejemplo en los linfocitos cebados con antígeno).
Requerimientos de contactos estrechos célula a célula: la citoquina sólo alcanza concentraciones adecuadas para actuar en el estrecho espacio que queda entre dos células interactuantes; recordar por ejemplo las "bolsas" que se forman en el conjugado TH:B, donde se alcanzan mejor esos niveles de citoquinas.
Corta vida media de las citoquinas en sangre y fluidos, lo que asegura que sólo van a actuar en un estrecho margen de tiempo, en las cercanías de la zona donde se produjeron.
14.4 RECEPTORES DE CITOQUINAS
14.4.1 Estructura general de las familias de receptores de citoquinas
Hay diversos tipos de receptores de membrana para citoquinas, pero se pueden agrupar en cinco familias:
Familia de receptores de citoquinas de la superfamilia de las inmunoglobulinas, que poseen varios dominios extracelulares de tipo Ig. Como ejemplo, el receptor específico para la IL-1.
Familia de clase I de receptores de citoquinas (=familia de receptores de hematopoyetinas). (Veremos más detalles de esta familia un poco más adelante).
Familia de clase II de receptores de citoquinas (=familia de receptores de interferones). Ejemplos de ligandos son los interferones no inmunes (IFN-a y b) y el IFN-g.
Familia de receptores de TNF: sus miembros se caracterizan por un dominio extracelular rico en cisteínas. Ejemplos de ligandos: TNF-a, TNF-b, CD40.
Familia de receptores de quimioquinas: son proteínas integrales de membrana, con 7 hélices a inmersas en la bicapa lipídica. Interaccionan, por el lado que da al citoplasma con proteínas de señalización triméricas que unen GTP. Ejemplos de quimioquinas que se unen a miembros de esta familia: IL-8, RANTES.
La mayor parte de los receptores de citoquinas del sistema inmune pertenecen a la familia de clase I (de receptores de hematopoyetinas). Todos sus miembros tienen en común poseer una proteína anclada a membrana, con un dominio extracelular en el que hay al menos un motivo característico llamado CCCC (cuatro cisteínas cercanas en posiciones equivalentes) y el llamado motivo WSXWS (Trp-Ser-X-Trp-Ser). (Adicionalmente, algunos miembros poseen dominios de tipo Ig y/o dominios de tipo fibronectina). Tras su porción transmembrana se encuentra una larga cola citoplásmica con ciertas tirosinas susceptibles de fosforilación.
La mayor parte de los receptores de clase I poseen dos proteínas de membrana:
Cadena a, que es la subunidad específica de la citoquina, sin capacidad de enviar señales al citoplasma;
Cadena b, una subunidad transductora de señal, que a menudo no es específica de citoquina, sino que es compartida por receptores de otras citoquinas. Esta cadena b es del tipo que hemos descrito más arriba (motivos CCCC y WSXWS).
La subunidad transductora de señal se necesita para formar el receptor de alta afinidad, y para transducir la señal al interior. Ello se logra porque tras la unión, se fosforilan ciertas tirosinas de la larga cola citoplásmica de la cadena transductora de señal.
En los esquemas comparativos de distintos receptores se puede apreciar que algunos de ellos poseen el mismo tipo de subunidad b . Este hecho permite explicar dos cualidades a las que ya hemos aludido: la redundancia y el antagonismo.
Por ejemplo: consideremos los receptores de IL-3, IL-5 y GM-CSF, que como se ve, comparten el mismo tipo (llamado KH97) de cadena b .
Redundancia: por separado, las tres citoquinas citadas, al tener sendos receptores que tienen el mismo tipo de cadena b , provocan los mismos efectos biológicos: proliferación de eosinófilos y desgranulación de basófilos.
Antagonismo: las tres citoquinas compiten entre sí por la unión de un número limitado de cadenas b con las a específicas de cada receptor.
La subfamilia de receptores a la que pertenece el receptor de IL-2 (el llamado IL2-R) consta de tres subunidades:
Cadena a específica de cada citoquina;
Dos subunidades (cadena b y cadena común gC) transductoras de señal.
Esta versión trimérica de IL-2R es la que une IL-2 con mayor afinidad, pero en realidad, el receptor para IL-2 no siempre aparece en esta forma, sino que existe una forma monomérica (IL-2Ra) de baja afinidad, sin cadenas transductoras, y por lo tanto incapaz de reenviar la señal, y una forma de afinidad intermedia (IL-2Rbg).
14.4.2 Transducción de señal
Recientemente se han producido avances importantes en el desentrañamiento de la ruta que conduce desde la unión de la citoquina con el receptor de la célula diana hasta la activación de la transcripción de los genes cuyos productos son responsables de los efectos de dichas citoquinas. He aquí un modelo general que se puede aplicar a muchos receptores de las clases I y II:
La citoquina provoca la dimerización de las dos subunidades del receptor (cadenas a y b), lo que coloca cercanas a sus respectivas colas citoplásmicas.
Una serie de proteín-quinasas de la familia de JAK (quinasas Jano) se unen a las colas agrupadas de las subunidades del receptor, con lo que se esas quinasas se activan.
Las JAK se autofosforilan.
Las JAK fosforilan a su vez determinadas tirosinas de las colas del receptor,
Entonces proteínas de otra familia, llamada STAT (iniciales inglesas de transductores de señal y activadores de transcripción) se unen a algunas de las tirosinas fosforiladas de las colas del receptor, quedando cerca de las JAK.
Las JAK fosforilan a las STAT unidas a las colas del receptor.
Al quedar fosforiladas, las STAT pierden su afinidad por las colas del receptor, y en cambio tienden a formar dímeros entre sí. (Las tirosinas fosforiladas que han quedado libres en las colas del receptor sirven para unir nuevos monómeros de STATs).
Los dímeros de STAT fosforilados emigran al núcleo de la célula, donde actúan ahora como activadores de la transcripción de ciertos genes, al unirse a secuencias especiales en la parte 5’ respecto de las respectivas porciones codificadoras.
14.5 ANTAGONISTAS DE CITOQUINAS
La actividad biológica de las citoquinas está regulada fisiológicamente por dos tipos de antagonistas:
los que provocan el bloqueo del receptor al unirse a éste:
los que inhiben la acción de la citoquina al unirse a ésta.
Como ejemplo de bloqueador de receptor tenemos el antagonista del receptor de IL-1 (IL-1Ra), que bloquea la unión de IL-1a o IL-1b . Desempeña un papel en la regulación de la intensidad de la respuesta inflamatoria. En la actualidad se está investigando su potencial clínico en el tratamiento de enfermedades que cursan con inflamación crónica.
Los inhibidores de citoquinas suelen ser versiones solubles de los respectivos receptores (y se suelen denominar anteponiendo una "s" al nombre del receptor): la rotura enzimática de la porción extracelular libera un fragmento soluble que retiene su capacidad de unirse a la citoquina. Existen ejemplos de versiones solubles de los receptores IL-2R, IL-4R, IL-7R, IFN-g R, TNF-a R, TNF-b R.
El mejor caracterizado es el sIL-2R (versión soluble del receptor de la interleuquina 2), que se libera durante la activación crónica de los linfocitos T, y que corresponde a los 192 aminoácidos N-terminales de la subunidad a . Este sIL-2R se puede unir a la IL-2, impidiendo su interacción con el auténtico receptor de membrana, con lo que esto supone un control sobre el exceso de activación de los linfocitos T. Este inhibidor se usa de hecho en clínica como un marcador de la existencia de activación crónica (caso, p. ej., de las enfermedades autoinmunes, rechazo de injertos y SIDA).
Algunos virus han evolucionado (como parte de sus mecanismos de evasión del sistema defensivo del hospedador) para producir proteínas que se unen e inactivan a las citoquinas.
Por ejemplo, los poxvirus codifican una proteína soluble que se une al TNF-a , y otra que se liga a la IL-1. Con ello logran reducir el alcance e intensidad de los mecanismos inflamatorios naturales que forman parte de la defensa del hospedador.
14.5 CONSECUENCIAS BIOLÓGICAS DE LA SECRECIÓN DE CITOQUINAS POR PARTE DE LOS LINFOCITOS TH1 Y TH2
En este apartado no vamos más que a recordar y profundizar un poco más en algo ya tratado en el contexto del tema 13: cómo el distinto espectro de citoquinas secretadas por las dos subpoblaciones de linfocitos TH determina distintos efectos biológicos, y cómo cada subpoblación controla a la otra.
Las células TH1 producen IL-2, IFN-g y TNF-b . Son responsables de funciones de inmunidad celular (activación de linfocitos TC e hipersensibilidad de tipo retardado), destinadas a responder a parásitos intracelulares (virus, protozoos, algunas bacterias).
Las células TH2 producen IL-4, IL-5, IL-10 e IL-13. Actúan como colaboradoras en la activación de las células B, y son más apropiadas para responder a bacterias extracelulares y a helmintos. También están implicadas en reacciones alérgicas (ya que la IL-4 activa la producción de IgE y la IL-5 activa a los eosinófilos).
En los años recientes está cada vez más claro que el resultado de la respuesta inmune depende en buena medida de los niveles relativos de células TH1 y TH2: en una respuesta a patógenos intracelulares existe un aumento de citoquinas de TH1, mientras que en respuestas alérgicas y ante helmintos es superior el nivel de las de TH2.
Un punto importante en todo esto es la existencia de una regulación cruzada entre TH1 y TH2:
El IFN-g secretado por las TH1 inhibe la proliferación de las TH2.
Por su lado, la IL-10 secretada por las TH2 inhibe la secreción de IL-2 e IFN-g por parte de las TH1. Esta inhibición en realidad no es directa: la IL-10 produce un descenso marcado de la cantidad de MHC-II de las células presentadoras de antígeno, que por lo tanto ya no pueden ejercer bien su papel de activar a las TH1. Además, las TH2 inhiben por sus citoquinas la producción en macrófagos del óxido nítrico (NO) y otros bactericidas, así como la secreción por estos macrófagos de IL-1, IL-6, IL-8 y otras citoquinas.
Este fenómeno de regulación negativa cruzada explica las ya antiguas observaciones de que existe una relación inversa entre la producción de anticuerpos y la hipersensibilidad de tipo retardado.
Obsérvese que los macrófagos y otras células presentadoras de antígeno también producen citoquinas (como la IL-12, descubierta hace relativamente poco tiempo) que regulan a su vez funciones inmunes efectoras. La IL-12 se produce en macrófagos activados en respuesta a infecciones bacterianas o de protozoos. Esta citoquina provoca la proliferación de células NK y TH1,que aumentan la producción de IFN-g . Este interferón inmune ayuda en la mayor activación de macrófagos. De esta forma se cierra este circuito de retrorregulación positiva entre macrófagos y TH1, destinado a potenciar funciones efectoras de la rama celular de la inmunidad.
Por otro lado, los macrófagos se ven inhibidos por IL-4 e IL-10 secretadas por los TH2 (de nuevo una manifestación de la inhibición cruzada entre la rama especializada en la respuesta humoral y la centrada en la respuesta celular ante parásitos intracelulares).
Otro aspecto que va quedando claro igualmente es que la predominancia de una u otra de las dos subpoblaciones de linfocitos TH depende a su vez del microambiente de citoquinas en que ocurriera la activación y maduración inicial a partir de linfocitos en reposo: por ejemplo, in vitro se ha visto que si un TH se activa por antígeno en presencia de IL-4, se desarrolla hasta TH2, mientras que si el entorno de activación es rico en IFN-g , se desarrolla hasta TH1.
Sus funciones son muy variadas, pero se pueden clasificar en unas pocas categorías:
diferenciación y maduración de células del sistema inmunitario;
comunicación entre células del sistema inmunitario;
en algunos casos, ejercen funciones efectoras directas.
En el pasado reciente hubo un cierto galimatías con la cuestión de su denominación. Así, muchas de las primeras citoquinas se descubrieron como señalizadoras entre leucocitos, por lo que se denominaron interleuquinas; otras eran secretadas por monocitos/macrófagos, por lo que se llamaron monoquinas. Sin embargo, muchas de esas sustancias son producidas por otros tipos celulares, por lo que se desaconseja el uso de esas denominaciones, para agruparlas a todas bajo el concepto de citoquinas. Las quimioquinas (o quimiocinas) son un tipo de citoquinas de pequeño tamaño, con papeles en la respuesta inflamatoria y la quimiotaxis de fagocitos.
14.2 PROPIEDADES GENERALES DE LAS CITOQUINAS
Las citoquinas son un grupo de proteínas secretadas de bajo peso molecular (por lo general menos de 30 kDa), producidas durante las respuestas inmunes natural y específica. Se unen a receptores específicos de la membrana de las células donde van a ejercer su función, iniciando una cascada de transducción intracelular de señal que altera el patrón de expresión génica, de modo que esas células diana producen una determinada respuesta biológica.
Las citoquinas son producidas por múltiples tipos celulares, principalmente del sistema inmune. Dentro del sistema inmune natural, los macrófagos son de las células más productoras de citoquinas, mientras que en el sistema específico lo son las células T colaboradoras.
La producción de las citoquinas suele ser breve (transitoria), limitada al lapso de tiempo que dura el estímulo (es decir, el agente extraño). En muchos casos ello se debe a que los correspondientes ARNm tienen una corta vida media, que a su vez depende de que las zonas 3’ no traducibles son ricas en A y U.
Considerando las diversas citoquinas, éstas pueden exhibir una o varias de las siguientes cualidades:
- pleiotropía (múltiples efectos al actuar sobre diferentes células).
- redundancia (varias citoquinas pueden ejercer el mismo efecto).
- sinergismo (dos o más citoquinas producen un efecto que se potencia mutuamente).
Por ejemplo, la acción conjunta de IL-4 e IL-5 induce en células B el cambio de clase para que produzcan IgE.
- antagonismo (inhibición o bloqueo mutuo de sus efectos).
Por ejemplo, el IFN-g bloquea el cambio de clase promovido por IL-4.
Las citoquinas ejercen su acción al unirse a receptores específicos para cada citoquina en la superficie de la célula en la que ejercen el efecto. La afinidad de cada receptor hacia su citoquina correspondiente suele ser bastante alta, del orden de lo femtomolar (10-15 M) a lo picomolar (10-12 M).
Utilizando la analogía de lo que ocurre con las hormonas del sistema endocrino, las acción de las citoquinas se puede clasificar en:
- de tipo autocrino
- de tipo paracrino
- (en pocas ocasiones) de tipo endocrino.
Las citoquinas "controlan" el sistema inmune de varias maneras, que podemos agrupar de la siguiente manera:
regulando (activando o inhibiendo) la activación, proliferación y diferenciación de varios tipos de células;
regulando la secreción de anticuerpos y de otras citoquinas.
14.3 ESTRUCTURA Y FUNCIÓN DE LAS CITOQUINAS
Las citoquinas son proteínas o glucoproteínas de menos de 30 kDa. Muchas de ellas pertenecen a la llamada familia de las hematopoyetinas, y tienen estructuras terciarias parecidas: una configuración a base de un conjunto de cuatro hélices a , con poca estructura en lámina b .
Generalmente actúan como mensajeros intercelulares que suelen intervenir en la maduración y amplificación de la respuesta inmune, provocando múltiples actividades biológicas una vez que se unen a los receptores específicos de las células diana adecuadas.
Aunque existen muchos tipos de células productoras citoquinas (ya hemos ido viendo unas cuantas en los temas anteriores), los más importantes son los linfocitos TH y los macrófagos, ya que sus citoquinas son esenciales para que se produzca la respuesta inmune una vez que se activan las células T y B por el contacto con las correspondientes células presentadoras de antígeno.
Principales tipos de respuesta mediatizados por la acción de las citoquinas:
activación de los mecanismos de inmunidad natural:
activación de los macrófagos y otros fagocitos
activación de las células NK
activación de los eosinófilos
inducción de las proteínas de fase aguda en el hígado (ver tema 16, sobre el complemento).
Activación y proliferación de células B, hasta su diferenciación a células plasmáticas secretoras de anticuerpos.
Intervención en la respuesta celular específica.
Intervención en la reacción de inflamación, tanto aguda como crónica.
Control de los procesos hematopoyéticos de la médula ósea.
Inducción de la curación de las heridas.
Hay una aparente paradoja de las citoquinas que debemos explicar: ¿Por qué las citoquinas, que son inespecíficas respecto del antígeno, pueden ejercer acciones de modo específico? Veamos varios mecanismos:
Regulación muy fina de los receptores de cada citoquina: los receptores celulares indispensables para que una citoquina ejerza su papel sólo se expresan en tipos celulares concretos una vez que éstos han interaccionado con el antígeno (pensemos por ejemplo en los linfocitos cebados con antígeno).
Requerimientos de contactos estrechos célula a célula: la citoquina sólo alcanza concentraciones adecuadas para actuar en el estrecho espacio que queda entre dos células interactuantes; recordar por ejemplo las "bolsas" que se forman en el conjugado TH:B, donde se alcanzan mejor esos niveles de citoquinas.
Corta vida media de las citoquinas en sangre y fluidos, lo que asegura que sólo van a actuar en un estrecho margen de tiempo, en las cercanías de la zona donde se produjeron.
14.4 RECEPTORES DE CITOQUINAS
14.4.1 Estructura general de las familias de receptores de citoquinas
Hay diversos tipos de receptores de membrana para citoquinas, pero se pueden agrupar en cinco familias:
Familia de receptores de citoquinas de la superfamilia de las inmunoglobulinas, que poseen varios dominios extracelulares de tipo Ig. Como ejemplo, el receptor específico para la IL-1.
Familia de clase I de receptores de citoquinas (=familia de receptores de hematopoyetinas). (Veremos más detalles de esta familia un poco más adelante).
Familia de clase II de receptores de citoquinas (=familia de receptores de interferones). Ejemplos de ligandos son los interferones no inmunes (IFN-a y b) y el IFN-g.
Familia de receptores de TNF: sus miembros se caracterizan por un dominio extracelular rico en cisteínas. Ejemplos de ligandos: TNF-a, TNF-b, CD40.
Familia de receptores de quimioquinas: son proteínas integrales de membrana, con 7 hélices a inmersas en la bicapa lipídica. Interaccionan, por el lado que da al citoplasma con proteínas de señalización triméricas que unen GTP. Ejemplos de quimioquinas que se unen a miembros de esta familia: IL-8, RANTES.
La mayor parte de los receptores de citoquinas del sistema inmune pertenecen a la familia de clase I (de receptores de hematopoyetinas). Todos sus miembros tienen en común poseer una proteína anclada a membrana, con un dominio extracelular en el que hay al menos un motivo característico llamado CCCC (cuatro cisteínas cercanas en posiciones equivalentes) y el llamado motivo WSXWS (Trp-Ser-X-Trp-Ser). (Adicionalmente, algunos miembros poseen dominios de tipo Ig y/o dominios de tipo fibronectina). Tras su porción transmembrana se encuentra una larga cola citoplásmica con ciertas tirosinas susceptibles de fosforilación.
La mayor parte de los receptores de clase I poseen dos proteínas de membrana:
Cadena a, que es la subunidad específica de la citoquina, sin capacidad de enviar señales al citoplasma;
Cadena b, una subunidad transductora de señal, que a menudo no es específica de citoquina, sino que es compartida por receptores de otras citoquinas. Esta cadena b es del tipo que hemos descrito más arriba (motivos CCCC y WSXWS).
La subunidad transductora de señal se necesita para formar el receptor de alta afinidad, y para transducir la señal al interior. Ello se logra porque tras la unión, se fosforilan ciertas tirosinas de la larga cola citoplásmica de la cadena transductora de señal.
En los esquemas comparativos de distintos receptores se puede apreciar que algunos de ellos poseen el mismo tipo de subunidad b . Este hecho permite explicar dos cualidades a las que ya hemos aludido: la redundancia y el antagonismo.
Por ejemplo: consideremos los receptores de IL-3, IL-5 y GM-CSF, que como se ve, comparten el mismo tipo (llamado KH97) de cadena b .
Redundancia: por separado, las tres citoquinas citadas, al tener sendos receptores que tienen el mismo tipo de cadena b , provocan los mismos efectos biológicos: proliferación de eosinófilos y desgranulación de basófilos.
Antagonismo: las tres citoquinas compiten entre sí por la unión de un número limitado de cadenas b con las a específicas de cada receptor.
La subfamilia de receptores a la que pertenece el receptor de IL-2 (el llamado IL2-R) consta de tres subunidades:
Cadena a específica de cada citoquina;
Dos subunidades (cadena b y cadena común gC) transductoras de señal.
Esta versión trimérica de IL-2R es la que une IL-2 con mayor afinidad, pero en realidad, el receptor para IL-2 no siempre aparece en esta forma, sino que existe una forma monomérica (IL-2Ra) de baja afinidad, sin cadenas transductoras, y por lo tanto incapaz de reenviar la señal, y una forma de afinidad intermedia (IL-2Rbg).
14.4.2 Transducción de señal
Recientemente se han producido avances importantes en el desentrañamiento de la ruta que conduce desde la unión de la citoquina con el receptor de la célula diana hasta la activación de la transcripción de los genes cuyos productos son responsables de los efectos de dichas citoquinas. He aquí un modelo general que se puede aplicar a muchos receptores de las clases I y II:
La citoquina provoca la dimerización de las dos subunidades del receptor (cadenas a y b), lo que coloca cercanas a sus respectivas colas citoplásmicas.
Una serie de proteín-quinasas de la familia de JAK (quinasas Jano) se unen a las colas agrupadas de las subunidades del receptor, con lo que se esas quinasas se activan.
Las JAK se autofosforilan.
Las JAK fosforilan a su vez determinadas tirosinas de las colas del receptor,
Entonces proteínas de otra familia, llamada STAT (iniciales inglesas de transductores de señal y activadores de transcripción) se unen a algunas de las tirosinas fosforiladas de las colas del receptor, quedando cerca de las JAK.
Las JAK fosforilan a las STAT unidas a las colas del receptor.
Al quedar fosforiladas, las STAT pierden su afinidad por las colas del receptor, y en cambio tienden a formar dímeros entre sí. (Las tirosinas fosforiladas que han quedado libres en las colas del receptor sirven para unir nuevos monómeros de STATs).
Los dímeros de STAT fosforilados emigran al núcleo de la célula, donde actúan ahora como activadores de la transcripción de ciertos genes, al unirse a secuencias especiales en la parte 5’ respecto de las respectivas porciones codificadoras.
14.5 ANTAGONISTAS DE CITOQUINAS
La actividad biológica de las citoquinas está regulada fisiológicamente por dos tipos de antagonistas:
los que provocan el bloqueo del receptor al unirse a éste:
los que inhiben la acción de la citoquina al unirse a ésta.
Como ejemplo de bloqueador de receptor tenemos el antagonista del receptor de IL-1 (IL-1Ra), que bloquea la unión de IL-1a o IL-1b . Desempeña un papel en la regulación de la intensidad de la respuesta inflamatoria. En la actualidad se está investigando su potencial clínico en el tratamiento de enfermedades que cursan con inflamación crónica.
Los inhibidores de citoquinas suelen ser versiones solubles de los respectivos receptores (y se suelen denominar anteponiendo una "s" al nombre del receptor): la rotura enzimática de la porción extracelular libera un fragmento soluble que retiene su capacidad de unirse a la citoquina. Existen ejemplos de versiones solubles de los receptores IL-2R, IL-4R, IL-7R, IFN-g R, TNF-a R, TNF-b R.
El mejor caracterizado es el sIL-2R (versión soluble del receptor de la interleuquina 2), que se libera durante la activación crónica de los linfocitos T, y que corresponde a los 192 aminoácidos N-terminales de la subunidad a . Este sIL-2R se puede unir a la IL-2, impidiendo su interacción con el auténtico receptor de membrana, con lo que esto supone un control sobre el exceso de activación de los linfocitos T. Este inhibidor se usa de hecho en clínica como un marcador de la existencia de activación crónica (caso, p. ej., de las enfermedades autoinmunes, rechazo de injertos y SIDA).
Algunos virus han evolucionado (como parte de sus mecanismos de evasión del sistema defensivo del hospedador) para producir proteínas que se unen e inactivan a las citoquinas.
Por ejemplo, los poxvirus codifican una proteína soluble que se une al TNF-a , y otra que se liga a la IL-1. Con ello logran reducir el alcance e intensidad de los mecanismos inflamatorios naturales que forman parte de la defensa del hospedador.
14.5 CONSECUENCIAS BIOLÓGICAS DE LA SECRECIÓN DE CITOQUINAS POR PARTE DE LOS LINFOCITOS TH1 Y TH2
En este apartado no vamos más que a recordar y profundizar un poco más en algo ya tratado en el contexto del tema 13: cómo el distinto espectro de citoquinas secretadas por las dos subpoblaciones de linfocitos TH determina distintos efectos biológicos, y cómo cada subpoblación controla a la otra.
Las células TH1 producen IL-2, IFN-g y TNF-b . Son responsables de funciones de inmunidad celular (activación de linfocitos TC e hipersensibilidad de tipo retardado), destinadas a responder a parásitos intracelulares (virus, protozoos, algunas bacterias).
Las células TH2 producen IL-4, IL-5, IL-10 e IL-13. Actúan como colaboradoras en la activación de las células B, y son más apropiadas para responder a bacterias extracelulares y a helmintos. También están implicadas en reacciones alérgicas (ya que la IL-4 activa la producción de IgE y la IL-5 activa a los eosinófilos).
En los años recientes está cada vez más claro que el resultado de la respuesta inmune depende en buena medida de los niveles relativos de células TH1 y TH2: en una respuesta a patógenos intracelulares existe un aumento de citoquinas de TH1, mientras que en respuestas alérgicas y ante helmintos es superior el nivel de las de TH2.
Un punto importante en todo esto es la existencia de una regulación cruzada entre TH1 y TH2:
El IFN-g secretado por las TH1 inhibe la proliferación de las TH2.
Por su lado, la IL-10 secretada por las TH2 inhibe la secreción de IL-2 e IFN-g por parte de las TH1. Esta inhibición en realidad no es directa: la IL-10 produce un descenso marcado de la cantidad de MHC-II de las células presentadoras de antígeno, que por lo tanto ya no pueden ejercer bien su papel de activar a las TH1. Además, las TH2 inhiben por sus citoquinas la producción en macrófagos del óxido nítrico (NO) y otros bactericidas, así como la secreción por estos macrófagos de IL-1, IL-6, IL-8 y otras citoquinas.
Este fenómeno de regulación negativa cruzada explica las ya antiguas observaciones de que existe una relación inversa entre la producción de anticuerpos y la hipersensibilidad de tipo retardado.
Obsérvese que los macrófagos y otras células presentadoras de antígeno también producen citoquinas (como la IL-12, descubierta hace relativamente poco tiempo) que regulan a su vez funciones inmunes efectoras. La IL-12 se produce en macrófagos activados en respuesta a infecciones bacterianas o de protozoos. Esta citoquina provoca la proliferación de células NK y TH1,que aumentan la producción de IFN-g . Este interferón inmune ayuda en la mayor activación de macrófagos. De esta forma se cierra este circuito de retrorregulación positiva entre macrófagos y TH1, destinado a potenciar funciones efectoras de la rama celular de la inmunidad.
Por otro lado, los macrófagos se ven inhibidos por IL-4 e IL-10 secretadas por los TH2 (de nuevo una manifestación de la inhibición cruzada entre la rama especializada en la respuesta humoral y la centrada en la respuesta celular ante parásitos intracelulares).
Otro aspecto que va quedando claro igualmente es que la predominancia de una u otra de las dos subpoblaciones de linfocitos TH depende a su vez del microambiente de citoquinas en que ocurriera la activación y maduración inicial a partir de linfocitos en reposo: por ejemplo, in vitro se ha visto que si un TH se activa por antígeno en presencia de IL-4, se desarrolla hasta TH2, mientras que si el entorno de activación es rico en IFN-g , se desarrolla hasta TH1.
Las citoquinas o (citosinas) son un grupo de proteínas de
bajo peso molecular que actúan mediando interacciones complejas entre células
de linfoides, células inflamatorias y células hematopoyéticas.
Sus
funciones son muy variadas, pero se pueden clasificar en unas pocas categorías
:
Diferenciación y maduración de células del sistema
inmunitario
Comunicación entre células del sistema inmunitario
En algunos casos ejercen funciones efectoras directas.
En el pasado reciente hubo un cierto galimatías con la
cuestión de su denominación .Así, muchas de las primeras citoquinas se
descubrieron como señalización entre leucocitos, por lo que se denominaron
interleuquinas; otras eran secretadas por monocitos/macrófagos, por lo que se
llamaron monoquinas.Sin embargo, muchas de esas sustancias son producidas por
otros tipos celulares, por lo que se desaconseja el uso d esas denominaciones,
para agruparlas a todas, bajo el concepto de citoquinas. Las quimioquinas o
(quimiocinas) son un tipo de citoquinas de pequeño tamaño, con papeles n la
respuesta inflamatoria y la quimiotaxis de fagocitos.
Propiedades generales de las
citoquinas
Las citoquinas son un
grupo de proteínas secretadas de bajo
peso molecular (por lo general menos de 30 kDa), producidas durante las
respuestas inmunes natural y específicas .Se unen a receptores específicos de
la membrana de las células donde van a ejercer su función, iniciando una
cascada de transducción intracelular de señal que altera el patrón de expresión
genética, de modo que esas células diana producen una determinada respuesta
biológica.
Las citoquinas son
producidas por múltiples tipos celulares, principalmente del sistema inmune.
Dentro del sistema inmune natural, los macrófagos son de las células más
productoras de citoquinas, mientras que en el sistema específicos lo son las T
colaboradoras.
La producción de las
citoquinas suele ser breve (transitoria), limitada al lapso de tiempo que dura
el estímulo(es decir, el agente extraño).En muchos casos ello se debe a que los
correspondientes ARNm tienen una corta vida media, que a su vez depende de que
las zonas 3’ no traducibles son ricas en A y U.
Considerando las
diversas citoquinas, etapas pueden exhibir una o varias de las siguientes
cualidades:
- Plagiotropa
(múltiples efectos al actuar sobre diferentes células).
-Redundancia (varias
citoquinas pueden ejercer el mismo efecto).
-Sinergismo (dos o más
citoquinas producen un efecto que se potencia mutuamente).
Por ejemplo la acción conjunta de IL-4
e IL-5 induce en células B el cambio de clase para que produzcan IgE.
-Antagonismo
(inhibición o bloqueo mutuo de sus efectos).
Por ejemplo el IFN-y
bloquea el cambio de clase promovido por IL-4.
Las citoquinas
ejercen su acción al unirse a receptores específicos para cada citoquinas en
superficie de la célula en la que ejercen el efecto. La afinidad de cada
receptor hacia su citoquina correspondiente suele ser bastante alta, del orden
de lo femtomolar (10-15M) a lo pico molar (10-12M).
Utilizando la
analogía de lo que ocurre con las hormonas del sistema endocrino, las acciones
de las citoquinas se puede clasificar en:
-de tipo autocrino
-de tipo paracrino
-en pocas ocasiones
de tipo endocrino
Las citoquinas
“controlan” el sistema de varias maneras, que podemos agrupar de la siguiente
manera:
Regulando(activando o
inhibiendo) la activación, proliferación y diferenciación de varios tipos d células.
Regulando la
secreción de anticuerpos y de otras citoquinas.
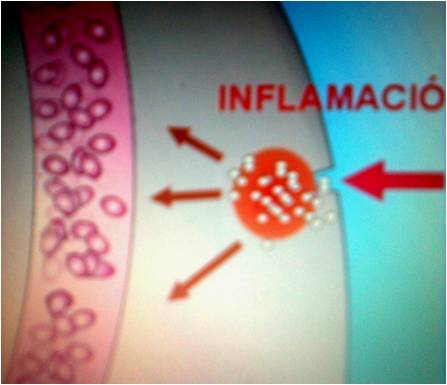
INFLAMACIÓN
La
inflamación es la respuesta, del sistema inmunológico de un organismo, al daño
causado a sus células y tejidos vascularizados por patógenos bacterianos y por
cualquier otro agresor de naturaleza biológica, química, física o mecánica.
Aunque dolorosa, la inflamación es, normalmente, una respuesta reparadora; un
proceso que implica un enorme gasto de energía metabólica. En ocasiones,
transcurre hacia una situación crónica que suele dar lugar a una enfermedad
degenerativa como artritisarteriosclerosis o, incluso, cáncer. Aunque suele
acompañarse de una respuesta generalizada «respuesta de fase aguda»
caracterizada por un cuadro clínico pasajero de sensación de malestar, fiebre y
modificación del perfil de las proteínas y leucocitos circulantes, en
ocasiones, la inflamación aguda local provoca una reacción orgánica
generalizada «síndrome de respuesta inflamatoria sistémica» que, en una
secuencia de reacciones a modo de espiral sin control «inflamación maligna»,
conduce al fracaso funcional de los diferentes órganos y sistemas «fracaso
multiorgánico» y, tras ello, a la muerte
del individuo
En
cualquier caso, la respuesta inflamatoria está coordinada por un copioso
dispositivo de mediadores que se organizan en complejas redes reguladoras. Para
disecar tal entramado, es útil agrupar esas señales en categorías funcionales y
distinguir entre inductores y mediadores de la respuesta inflamatoria. Los
primeros son señales que inician el proceso; activan sensores especializados
que suscitan la producción de lotes específicos de mediadores. Estos, a su vez,
alteran los estados funcionales de células, tejidos y órganos que son los
efectores de la inflamación, de manera que permitan su adaptación primero y reparación
después, del daño infringido por el inductor
I. BOSQUEJO
HISTÓRICO Para comprender la respuesta
inflamatoria, tal como hoy se entiende, es oportuno repasar las raíces de
nuestro conocimiento sobre tal fenómeno. La perspectiva histórica indica que el
progreso científico no es continuo, y que la ciencia está influida por la
personalidad de quienes la hacen avanzar. La historia de la investigación sobre
la inflamación se extiende más allá de dos mil años; de ellos, doscientos de
investigación en el contexto celular y, los veinte últimos, en el entorno
molecular.
Los signos
y síntomas inflamatorios se conocen desde antiguo. Los egipcios describieron
abscesos y úlceras, y el Código de Hammurabi contiene instrucciones para tratar
abscesos oculares . Sin embargo, fue el médico griego Hipócrates de Cos4 quién
introdujo las palabras edema y erisipela para describir la inflamación.
Hipócrates consideró la inflamación como el inicio de un proceso de resolución
o de curación. La primera descripción comprensiva de la inflamación se
encuentra en los escritos de Aulo C. Celso , quién no fue médico ni científico.
En su De Medicina introdujo los que consideran los síntomas cardinales de la
inflamación: rubor, tumor, color y dolor. Galeno de Pérgamo6 añadió un quinto
signo: functio laesa. Galeno, que introdujo el concepto de los cuatro humores
vitales (sanguis, pituita, chole y melaine chole), consideró la inflamación
como un desajuste en la relación de esos humores . No consideró que el pus
fuera perjudicial (pus laudable), aunque debía facilitarse su evacuación
mediante punción. Galeno advirtió del efecto perjudicial del frío sobre un
edema inflamatorio, porque el frío tornaba la lividez del tejido en una scirros
(cicatriz patológica), y pensó que la sangre percolaba a través de las paredes
de las arterias hacia el tejido.
Con todo,
la investigación del proceso inflamatorio no comenzó hasta la invención del
microscopio compuesto por el científico holandés Zacharias Janssen (1580-1638),
alrededor del año 1595. Antoni van Leeuwenhoek (1632-1723) construyó, en 1719,
el primer microscopio con la suficiente resolución óptica para observar
hematíes individuales moviéndose a través de los pequeños vasos sanguíneos,
aunque los leucocitos no fueron objeto de su interés. El médico Hermann
Boerhaave (1668-1738) utilizó el microscopio para observar vasos sanguíneos en
tejidos inflamados; concluyó que los vasos sanguíneos más pequeños eran
demasiado angostos para facilitar el flujo de sangre en el área inflamada, con
lo se generaba calor debido a la fricción. Un discípulo de Boerhave, Hieronymus
D. Gaubius (1705-1780), encontró que la inflamación promovía las condiciones
para que coagulara la sangre, y en 1796 apareció el texto “Treatise of the
blood, inflammation, and gun-shot wounds”, obra póstuma del inglés John Hunter
(1728-1793).
En el año
18248 , que corpúsculos sanguíneos individuales podían escapar a través de la
pared de los vasos y moverse con lentitud en la porción clara (de la
preparación), donde la velocidad de movimiento era muy lenta; ello en marcado
contraste con el torrente sanguíneo del que procedía la célula. Especuló sobre
la naturaleza de la trasmigración de los leucocitos, y sugirió que las paredes
de los vasos presentaban orificios que permitían a los elementos formes de la
sangreentrarenlos tejidos. Rudolf Wagner (1805-1864) tiene la primicia de la
descripción, en el año 1839 , del rodamiento leucocitario: «En el espacio claro
y brillante entre el torrente sanguíneo y la pared del vaso, que está rodeada
por varias fibras paralelas, puede observarse como los leucocitos se mueven
lentamente». Esta observación fue confirmada por Rudolf L. C. Virchow
(1821-1902) quién, en el año 187110, hizo notar que los leucocitos rodantes
podían adherirse transitoriamente a la pared del vaso y, en ocasiones, reentrar
en el torrente circulatorio. Observó la trasmigración leucocitaria, pero
atribuyó un papel nutritivo más que una función inflamatoria a este fenómeno.
Observaciones similares fueron hechas por William Addison (1802-1881) y por
Augustus V. Waller (1814- 1870), quienes ofrecieron, en la década de los años
1840, una clara descripción de la trasmigración de los leucocitos y de la
diapédesis de los eritrocitos en la inflamación secundaria a traumatismos, y
descubrieron que el pus estaba formado por leucocitos.-
Fagocitosis
Las
primeras células que atraviesan la pared vascular son los leucocitos
polimorfonucleares seguidos por los monocitos. Cuando los monocitos migran
hacia los tejidos son denominados macrófagos. En el proceso inflamatorio ambos
tipos de células funcionan como fagocitos.

La
fagocitosis es la ingestión de microorganismos o partículas de materia
resultantes de la rotura del tejido por los leucocitos polimorfonucleares,
monocitos o fagocitos hísticos. El proceso de fagocitosis incluye los procesos
de opsonización (recubrimiento de inmunoglobulinas o complemento a las
bacterias o antígenos), ingestión (después de la fijación la bacteria es
incorporada a la célula, rodeándola con seudópodos del fagocito), y por último
destrucción de las bacterias o antígenos (degradación por enzimas, pH ácido en
la vacuola, proteínas catiónicas, lactoferrinas, anión superóxido o peróxido de
hidrógeno).
Cuando las bacterias o los antígenos agresores
son destruidos, puede comenzar el proceso de la curación.
En
este proceso, una célula se deforma su membrana para formar un pequeño cono
alrededor de la pieza de material que va a ser absorbida, y entonces cierra los
lados del cono, que abraza la partícula en la membrana de la célula para crear
lo que se conoce como un fagosoma o vacuola de alimentos, como un pequeño sobre
de material rodeado por la membrana celular. El fagosoma, a su vez, se pasa a
la celda para la absorción por los lisosomas, las estructuras de las células
que se especializan en los materiales que entran en la célula de digerir. Los
lisosomas rompen el fagosoma abajo en sus materiales componentes, pasando
compuestos útiles a otras estructuras en la célula y expeler el resto como
material de desecho. En el caso de algunos materiales infecciosos o nocivos, el
fagosoma puede entrar en un peroxisomas, una estructura celular especial que
ayuda a liberar el cuerpo de toxinas.
En los
organismos unicelulares, la fagocitosis es una función crítica, ya que, sin
ella, el organismo no va a sobrevivir. Algunos de estos organismos se han
adaptado las características especiales que les permiten realizar un
seguimiento de los alimentos, orientándose en la dirección de las partículas
útiles que pueden ingerir. En los organismos multicelulares, que tiende a ser
un proceso más pasivo, pero sigue siendo crucial para la supervivencia de las
células individuales, asegurándose de que obtengan los nutrientes que necesitan
para funcionar.
Del
sistema inmune las células también realizan fagocitosis, atrapando a materiales
dañinos cuando entran en el cuerpo y destruirlos para que no puedan causar
daños. En algunos casos, el cuerpo puede responder con la inflamación, como una
inundación de células del sistema inmune se precipita a una ubicación para
hacer frente a invasores no deseados. Cuando el sistema inmunológico no puede
hacer frente a materiales dañinos, o cuando estos materiales resisten el
proceso de ingestión, las consecuencias para el organismo huésped pueden ser
bastante desagradable.
¿Qué funciones tiene?
En muchos organismos
superiores, la fagocitosis es tanto un medio de defensa ante microorganismos
invasores como de eliminación e incluso el reciclaje de tejidos muertos, además
es como una forma de nutrición para las células que realizan ésta función.
Células fagocitarias.
La fagocitosis se lleva a
cabo en células especializadas llamadas fagocitos, varias células en el cuerpo
humano ejercen funciones fagocitarias principalmente las células blancas como:
Neutrófilos.
Eosinofilos.
Basófilos.



Monocitos. Macrofagos.


ETAPAS DE LA FAGOCITOSIS
Comprende:
-Quimiotaxis -Adherencia -Ingestión -Digestión -Excreción

QUIMIOTAXIS
La Quimiotaxis se
presenta como un proceso fisiológico en donde el glóbulo blanco combate las
sustancias patógenas que han producido inflamación, este glóbulo se margina del
flujo sanguíneo, que en estas zonas de inflamación es turbulento, luego se
adhiere a la pared del vaso y transmigra a través de este para llegar a los
entes patógenos para fagocitarlos. Este proceso es considerado desde los
fenómenos de transporte electroquímico, flujos eléctricos y de concentración,
entre otros.



ADHERENCIA
Otros receptores sobre la membrana de los
leucocitos y otros fagocitos actúan como mecanismos de adherencia sobre los
microorganismos, sea a productos microbianos específicos o sobre opsoninas del
sistema inmune del hospedador como:
-Receptor de manosa. Este receptor tiene afinidad por los componentes de manosa presentes en las glucoproteínas y glucolípidos de las paredes celulares microbianos.
-Scavenger. Estos receptores se unen directamente a microorganismos y a moléculas de LDL modificadas.
-CD14. Es un ligando con preferencia específica al lipopolisacárido presente en ciertas bacterias y está asociado a un receptor tipo Toll.
-Receptor de manosa. Este receptor tiene afinidad por los componentes de manosa presentes en las glucoproteínas y glucolípidos de las paredes celulares microbianos.
-Scavenger. Estos receptores se unen directamente a microorganismos y a moléculas de LDL modificadas.
-CD14. Es un ligando con preferencia específica al lipopolisacárido presente en ciertas bacterias y está asociado a un receptor tipo Toll.

INGESTIÓN
La unión a receptores de adherencia promueve
señales de comunicación intracelular que resultan en la invaginación de la
membrana del fagocito rodeando al receptor y su ligando patogénico. Al rodear
por completo al complejo receptor: molécula, la membrana se une en sus extremos
y libera al interior de la célula un fagosoma. Esto puede ocurrir en más de un
punto de la membrana celular.

DIGESTIÓN
Una vez que el fagosoma está en el citoplasma
comienza la desintegración del mismo, proceso que se realiza por mecanismos
dependientes o independientes de Oxígeno. El primero se da tras activarse rutas
metabólicas que consumen oxígeno, lo cual produce la liberación de radicales libres
del oxígeno, que son tóxicos para los microorganismos. En el segundo caso es
donde intervienen los lisosomas, los cuales se unen al fagosoma conformando un
fagolisosoma, y liberando enzimas hidrolíticas que destruirán al antígeno.

CONCLUSION
En este trabajo ya elaborado pudimos aprender, conocer y ver
cada uno de estos tres componentes ; funciones, mecanismos y modos de acción
cave destacar que es de suma importancia manejar tanto los conceptos como
también se presentan y desarrollan cada uno de estos.
BIBLIOGRAFIA
--INMUNOLOGIA DE ROITT
--GRANADA.
--REV.R.ACAD.CIENC.EXACT.FISS.NAT.(ESP) VOL.102, NO. 1 PP
91-159
No hay comentarios.:
Publicar un comentario